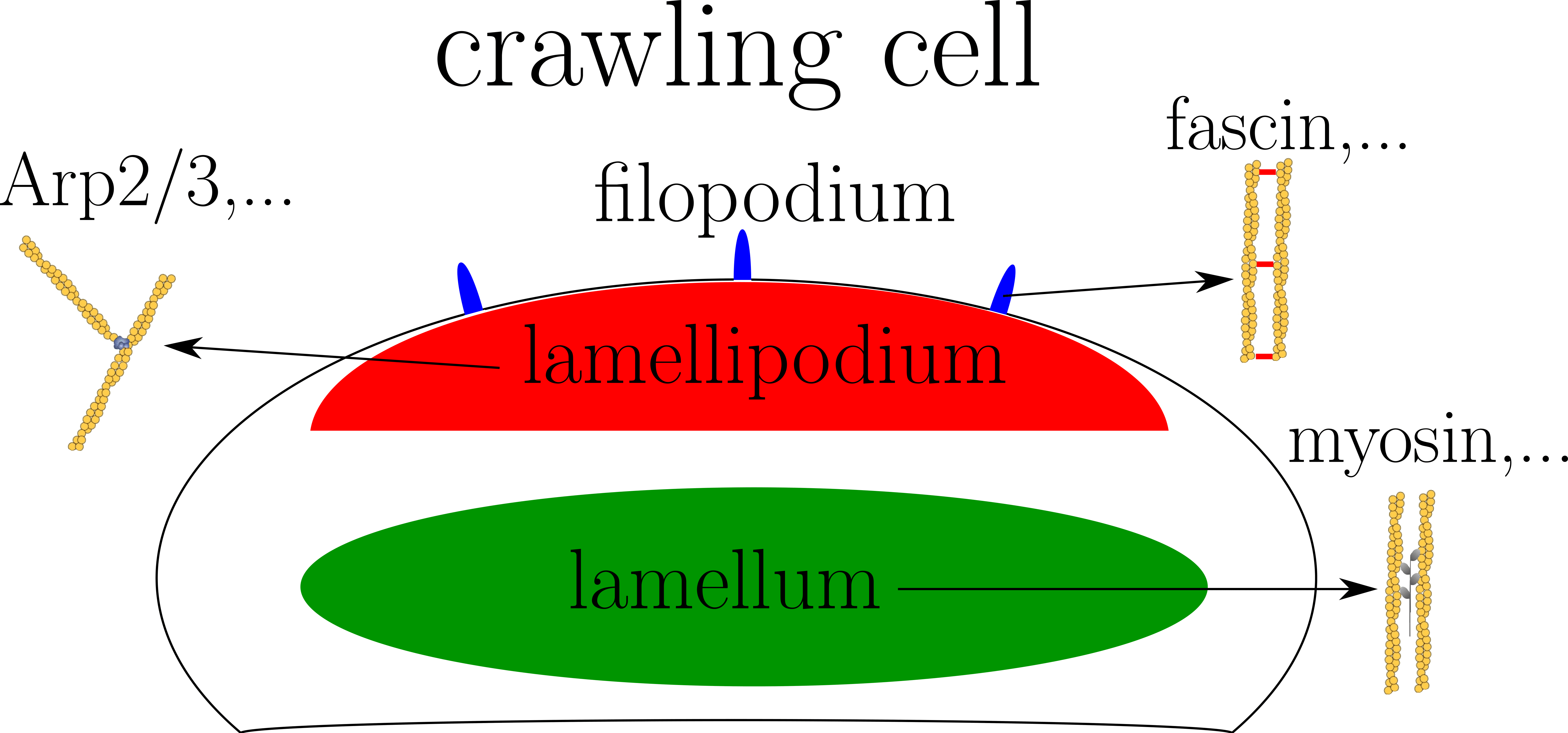
Cytoskeleton is a dynamical and complex structure involved in different processes like cell division or cell motility. One of its main ingredient is the actin biopolymer. The actin network is able to organize into different structures depending on the cellular state and function. For example, crawling cells has a lamellipodium formed by a scaffolded actin network at its front, a lamellum made of contractile mix of actin and myosin, and some filopodia composed of parallel actin filaments at the front (see figure on the right). Each struture has a specific location in the cell and a particular composition of actin binding proteins (ABPs). Nevertheless, the molecular principles driving the spatial segregation of these ABPs are still poorly understood. Many evidences show that some of ABPs bind cooperatively to actin. In addition, competition for adhesion exists between different families of ABPs. These two phenomenon could be the origin of the spatial segregation of ABPs. To answer this question I model both cooperative and competitive adhesion in Monte Carlo simulations.
Several studies model actin network as a continuum medium. This approximation is reasonable at cell scales. We know for example that a cellular extract of actomyosin (mix of actin and myosin) has a viscoelastic behaviour. I use a continuous approach to study dynamics of actin network in different situations. First, experiments show that after a complete depolymerization of actin in some cells, spots of actin appear and then waves emerge during reorganization of actin cytoskeleton. Using a continuous description of actin network I study the emergence of these waves and their influence on cell shape and motility. Then, some observations suggest that cell motility can be lead only by contraction of actomyosin at its rear. Modelling actomyosin as an active gel (viscous fluid with an additional active stress), I study motion of droplets enclosing this fluid (simulation picture on the right).
Lipid membranes are ubiquitous in biological systems. They are found in cell membrane, skin and articulations. These lipid membranes can experience a double-well potential leading to confinement. In cell adhesion, lipid membranes bind to a substrate while attached to the cytoskeleton. Double-well potentials has been measured in experiments mimicking this situation. Moreover, double-well potential can arise in cell-cell adhesion with two ligand-receptor pairs of different lengths. Starting from Helfrich model for lipid membranes, I study theoretically the adhesion dynamics of confined membranes (simulation result in figure on the right). In addition to confinement, lipid membranes can be sheared like in biolubrication systems for example (articulations,...). Some experiments suggest that lipid membranes could be the origin of a decrease in friction in these systems. Adding shear to the system, I both study membrane dynamics and friction forces.
 In collaboration with : Adrien Antkowiak and Alphée Michelot
In collaboration with : Adrien Antkowiak and Alphée Michelot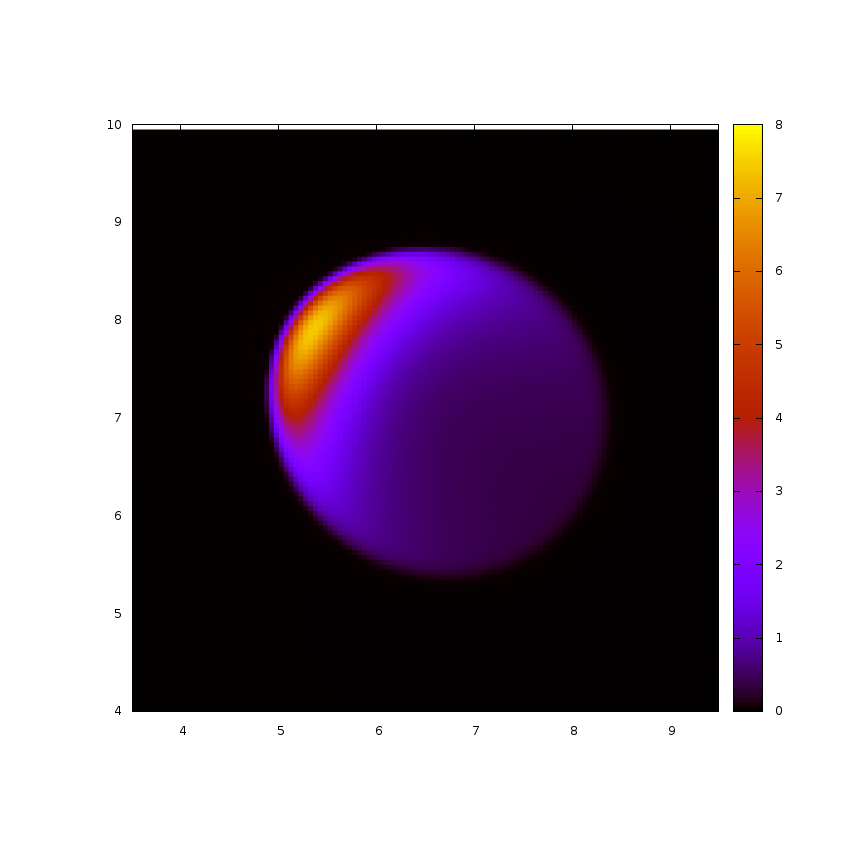 In collaboration with : Benno Liebchen and Davide Marenduzzo
In collaboration with : Benno Liebchen and Davide Marenduzzo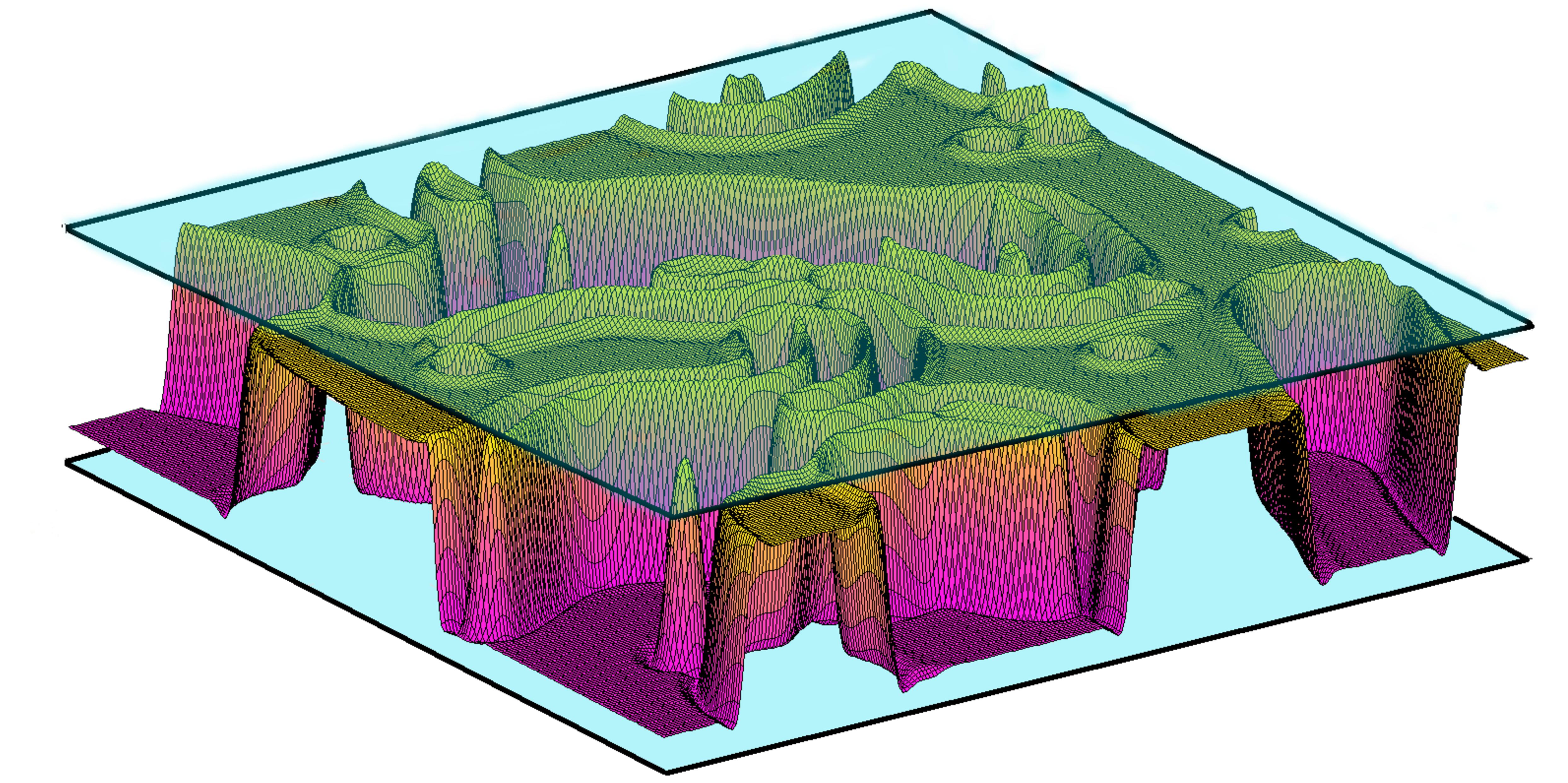 In collaboration with : Tung B.T. To, Paolo Politi and Olivier Pierre-Louis
In collaboration with : Tung B.T. To, Paolo Politi and Olivier Pierre-Louis