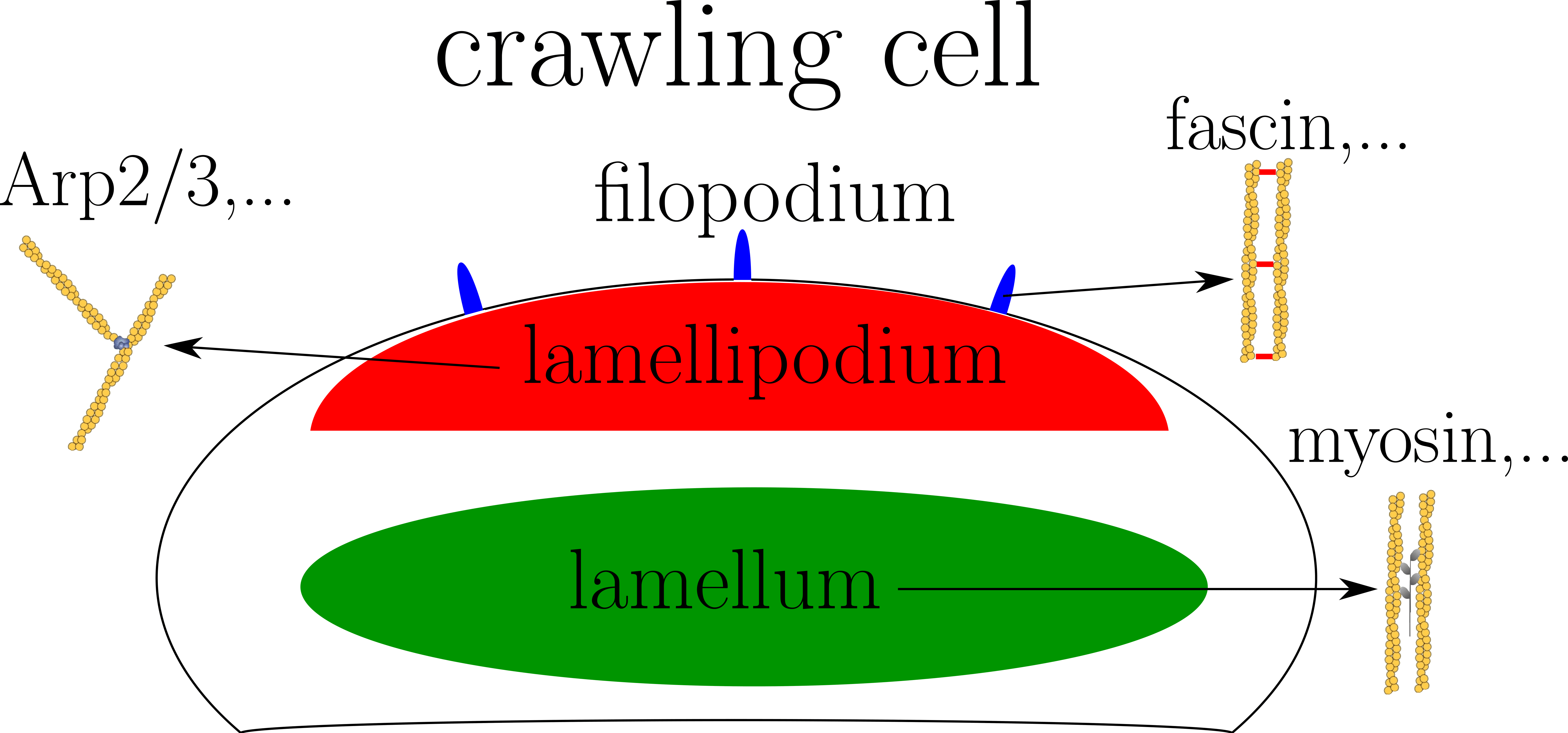
Le cytosquelette est une structure dynamique et complexe jouant un rôle dans plusieurs processus comme la division ou la motilité cellulaire. Un de ses principaux composants est le biopolymère d'actine. Le réseau d'actine peut s'organiser en plusieurs structures en fonction de l'état cellulaire et de sa fonction. Par exemple, une cellule migrante est composée d'un lamellipodium formé par un réseau branché d'actine à l'avant, d'un lamellum constitué d'un mélange contractile d'actine et de myosin, et de quelques filopodes faits de filaments d'actine parallèles (voir le schéma à droite). Chaque structure a une postion spécifique dans la cellule et une composition en protéines de liaison à l'actine (PLAs) particulière. Cependant, les principes moléculaires à l'origine de la ségrégation spatiale des PLAs sont encore mal compris. Beaucoup d'observations montrent que les PLAs se lient à l'actine de manière coopérative. De plus, une competition pour l'adhéions entre différentes familles de PLAs existe. Ces deux phénomènes pourrait être à l'origine de la ségrégation spatiale des PLAs. Pour apporter des éléments de réponse, je modélise l'adhésion à la fois coopérative et compétitive des PLAs dans des simulations de type Monte Carlo.
Plusieurs études modélisent le réseau d'actine comme un milieu continu, une approximation raisonnable à l'échelle de la cellule. Nous savons par exemple, qu'un extrait cellulaire d'actomyosine (mélange d'actine et de myosine) a un comportement viscoélastique. J'utilise une approche continue afin d'étudier la dynamique du réseau d'actine dans plusieurs situations. Tout d'abord, des expériences montrent qu'à la suite d'une dépolymérisation total de l'actine dans certaines cellules, des spots d'actine apparaissent puis des vagues émergent durant la réorganisation du cytosquelette. En utilisant une description continue du réseau d'actine, j'étudie l'émergence de ces vagues et leur influence sur la forme des cellules ainsi que leur motilité. Ensuite, des observations suggèrent que la motilité cellulaire peut être induite par la contraction seule de l'actomyosine à l'arrière de la cellule. En modélisant l'actomyosine comme un gel actif (fluide visqueux avec ajout d'un stress actif), j'étudie le mouvement de gouttes renfermant le fluide model (image tirée de simulations sur la droite).
Les membranes lipidiques sont omniprésentes dans les systèmes biologiques. On les trouve dans la membrane cellulaire, la peau et les articulations. Ces membranes lipidiques peuvent subir un potentiel double-pluits menant à un confinement. Dans l'adhésion cellulaire, les membranes lipidiques se lient à un substrat tout en restant attaché au cytosquelette. Des potentiels double-puits ont été mesurés dans des expériences reproduisant ce type de système. De plus, un potentiel double-puits peut émerger lors de l'adhésion de deux cellules entre elles, lorsqu'elles se lient via deux paires ligand-récepteur de différentes tailles. En partant du modèle de Helfrich pour les membranes lipidiques, j'étudie théoriquement la dynamique d'adhésion des membranes confinées (résultat de simulation sur la droite). En plus du confinement, les membranes lipidiques peuvent être cisaillées comme dans les systèmes de biolubrification par exemple (articulations,...). Des expériences suggèrent que les membranes lipidiques pourraient l'origine d'une baisse de friction dans ces systèmes. En rajoutant le ciasillement au système, j'étudie à la fois la dynamique de la membrane et les forces de friction.
 En collaboration avec : Adrien Antkowiak et Alphée Michelot
En collaboration avec : Adrien Antkowiak et Alphée Michelot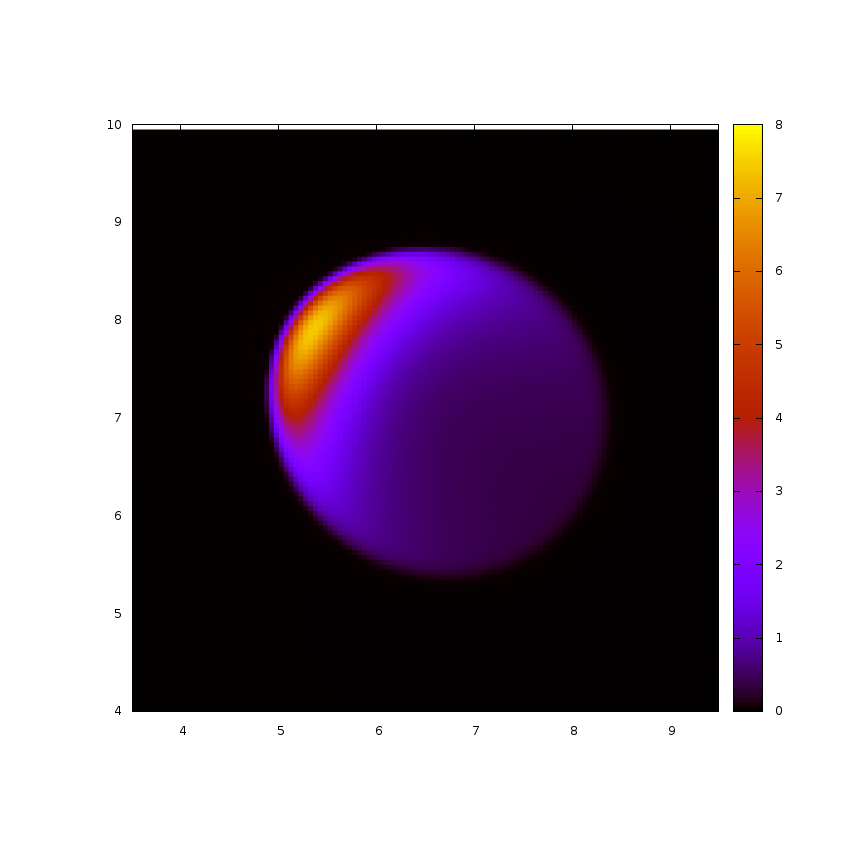 En collaboration avec : Benno Liebchen et Davide Marenduzzo
En collaboration avec : Benno Liebchen et Davide Marenduzzo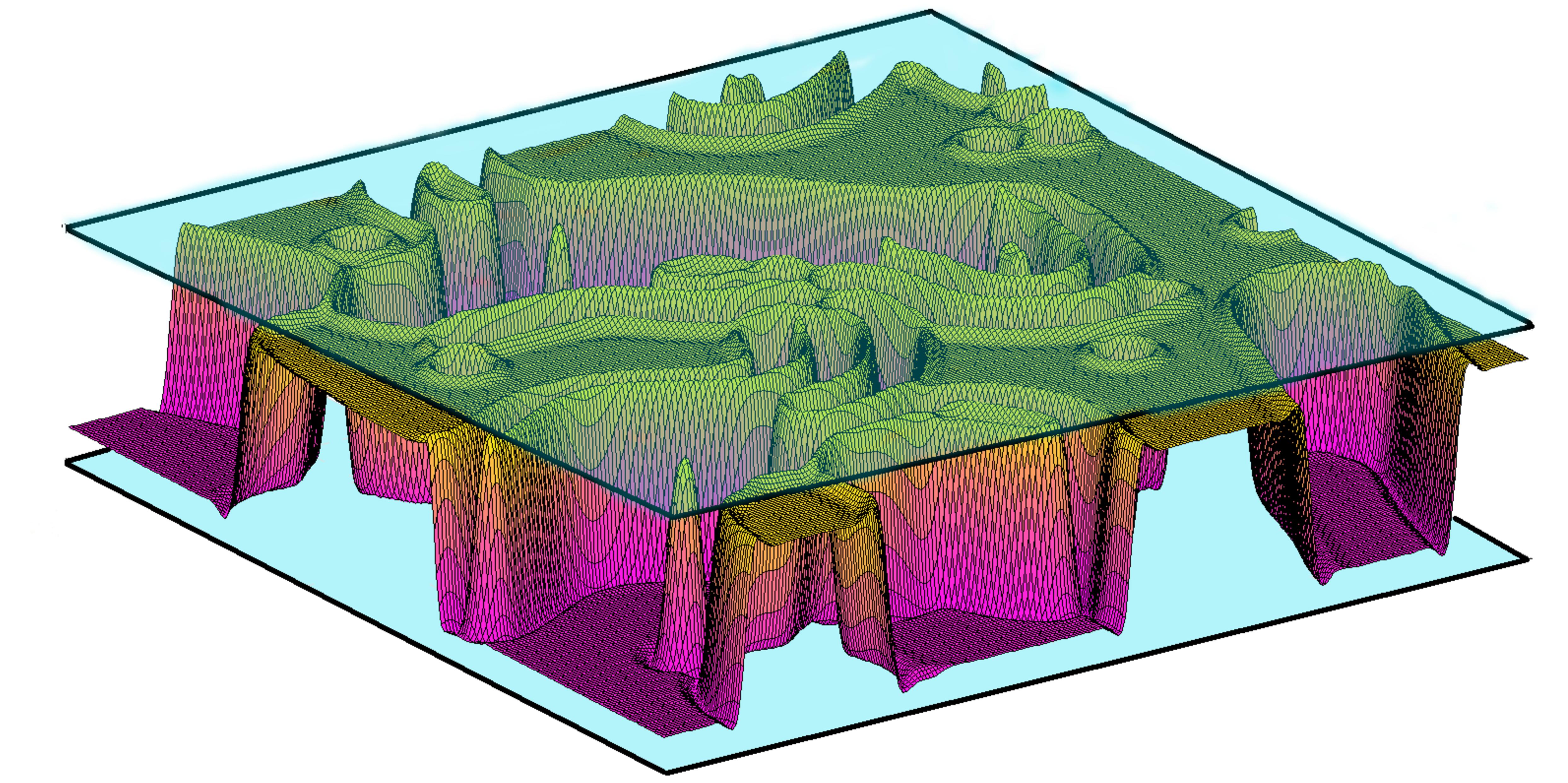 En collaboration avec : Tung B.T. To, Paolo Politi et Olivier Pierre-Louis
En collaboration avec : Tung B.T. To, Paolo Politi et Olivier Pierre-Louis